研究热点高分文献解读
近几年,外泌体,circRNA,m6A修饰和抗PD-1耐受已成为热门研究方向,获批国家自然科学基金的项目数显示出逐年递增。基于此,笔者今天给大家分享一篇高分文献来自Molecular Cancer (IF:41.44),基本涵盖了以上热点,并且在本文的肝癌细胞、CD8+ T细胞的研究模型下,阐述了这几者之间的关系,为大家后续研究提供参考。
肝细胞癌是一种高致死率的肿瘤,通常在晚期才被发现。近年来,免疫检查点阻断(immune checkpoint blocking, ICB)疗法,特别是针对PD1 (programmed cell death-1)/PD-L1 (programmed cell death ligand 1)的抗体,被广泛应用于癌症,大大延长了癌症患者的生存时间。PD1抗体已被批准用于晚期HCC的二线治疗。然而,只有17-18%的晚期肝癌患者对抗PD1抗体治疗达到完全治愈或部分缓解。因此,迫切需要探索抗PD1治疗的耐受性机制,并确定联合治疗策略,以提高其在肝癌治疗中的有效性。
肿瘤微环境(tumor microenvironment, TME)中CD8+T细胞功能失调导致的抗肿瘤免疫功能受损是癌症的一个特征。长期存在的肿瘤抗原和抑制性肿瘤微环境使抗肿瘤效应CD8+T细胞进入一种“T细胞耗竭”的功能失调状态。耗竭的CD8 + T细胞高度表达抑制受体,如PD1、T细胞免疫球蛋白和粘蛋白结构域-3 (TIM3)、淋巴细胞活化基因3 (LAG3)和含Ig和ITIM结构域的T细胞免疫受体(TIGIT)。此外,耗竭的CD8 + T细胞出现增殖能力下降,细胞毒功能减弱,释放效应细胞因子的能力减弱。
环状RNA (circRNAs)是一种具有闭环结构的非编码RNA,通过充当miRNA的海绵或与蛋白质结合来调节生物过程。许多circRNA参与了肝癌细胞的增殖、分化、凋亡、侵袭、干性和耐药过程。据报道,circRNA也被包装到外泌体中,并与肝癌的恶性发展过程和免疫抑制有关。然而,肝癌来源的外泌体环状RNA在免疫抑制微环境中的潜在功能和调控机制,特别是其对CD8 + T细胞的作用,仍需要更多的探索。
N6 -甲基腺苷(m6A)修饰是真核RNA的主要转录后修饰之一。m6A基因的失调与肝癌的癌变和进展有关。近年来,越来越多的研究表明m6A修饰的环状RNA在肝癌中发挥重要作用。例如,m6A介导的circMDK上调可促进肝癌发生;m6A修饰的circRNA-SORE维持肝癌中索拉非尼耐药;FTO介导的circGPR137B的m6A去甲基化抑制HCC的肿瘤发生和转移。然而,m6A修饰的环状RNA在调节肝癌抗肿瘤免疫中的作用仍需进一步研究。
Molecular Cancer
IF:41.44
Exosome-derived circCCAR1promotes CD8 + T-cell dysfunction and anti-PD1 resistance in hepatocellular carcinoma
背景:环状RNA (circRNAs)可以被封装到外泌体中参与细胞间的通讯,影响多种肿瘤恶性发展。CD8 + T细胞功能障碍是介导肝细胞癌(HCC)免疫逃逸的主要因素。然而,外泌体来源的环状RNA对 CD8 + T细胞功能障碍还有待进一步探讨。
方法:采用体外和体内两种方法观察circCCAR1对肝癌发生转移的影响功能实验。酶联法检测circCCAR1在CD8 + T 细胞功能障碍中的作用免疫吸附试验(ELISA)、western blotting和流式细胞术。利用CHIP,RNA pulldown、RIP等方法来探讨其作用机制,并且建立了重组人免疫系统组分小鼠模型(huNSG小鼠),探讨其在人体免疫系统中的作用,以及外泌体circCCAR1在肝癌抗PD1治疗耐药中的作用。
结果:肝癌患者的肿瘤组织和血浆外泌体中存在circCCAR1水平升高的现象。circCCAR1在体外和体内均能加速肝癌的生长和转移。E1A结合蛋白p300 (EP300)和真核翻译起始因子4A3 (EIF4A3)促进WTAP介导的m6A修饰,形成一个由circCCAR1/miR-127-5p/WTAP轴组成的反馈回路。circCCAR1是由肝癌细胞以异质核糖核蛋白A2/B1 (hnRNPA2B1)依赖性分泌的方式。外泌体circCCAR1被CD8 + T 细胞吸收,并通过稳定CD8 + T细胞功能紊乱PD-1蛋白质。circCCAR1促进了抗PD1免疫治疗的耐药性。此外,CCAR1与β-catenin蛋白的结合进一步增强了PD-L1的转录。
结论:在本项研究中,作者首先确定了一个新的研究目标circ_0000240,发现circCCAR1 /miR-127-5p/ WTAP通路促进肿瘤生长和转移肝细胞癌。肝癌细胞释放外泌体来源的circCCAR1,通过阻止PD1降解来促进CD8+T细胞功能障碍,从而增强肝癌细胞抗PD1治疗的耐药性。因此,本文结果揭示了circCCAR1在肝癌发展中发挥作用,并为研究环状RNA在免疫治疗肝癌中的功能奠定基础。
结论1
肝癌患者的肿瘤组织和血浆外泌体中存在circCCAR1水平升高的现象。
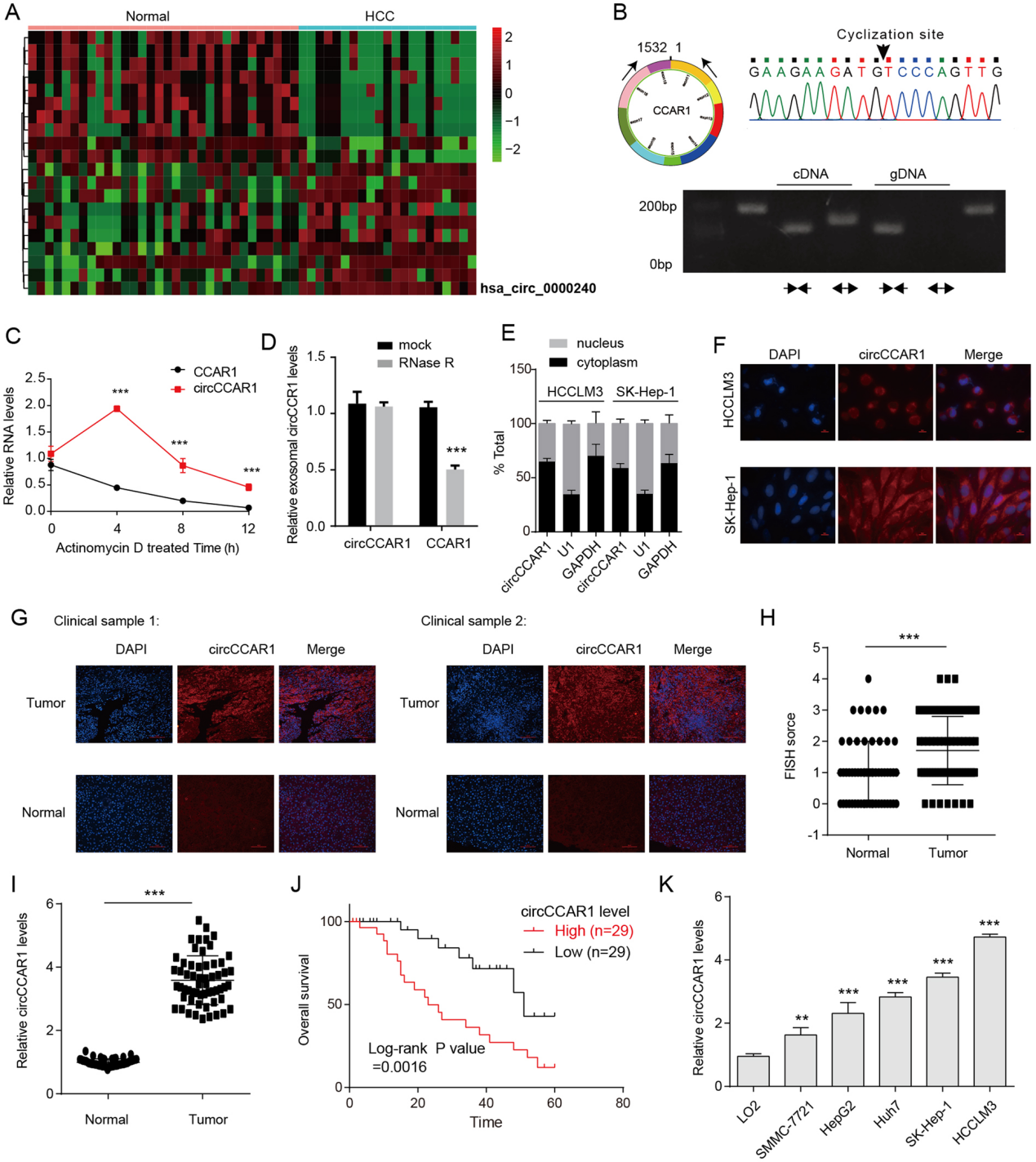
图1. A.聚类热图显示了健康受试者和HCC患者血清外泌体中的异常环状RNA。B.采用不同引物PCR法从HCCLM3细胞的cDNA或基因组DNA (gDNA)中扩增circCCAR1。连接序列经Sanger测序验证。C.放线菌素D处理后HCCLM3细胞中circCCAR1和CCAR1 mRNA的稳定性对比。D. RNase R消化后HCCLM3细胞CCAR1 mRNA和circCCAR1水平。E .通过细胞核质分离实验评估HCC细胞中circCCAR1的细胞内位置。F.采用FISH检测肝癌细胞中circCCAR1的细胞内定位。G.通过FISH获得HCC组织及邻近正常组织中circCCAR1的代表性荧光图像。H. FISH的统计数据。I.采用qRT-PCR法检测肝癌组织和正常组织中circCCAR1的表达水平。J.肝癌患者circCCAR1表达与总生存率的关系。K .检测LO2细胞和HCC细胞系中circCCAR1的表达水平。
结论2
circCCAR1在体外和体内促进HCC生长。
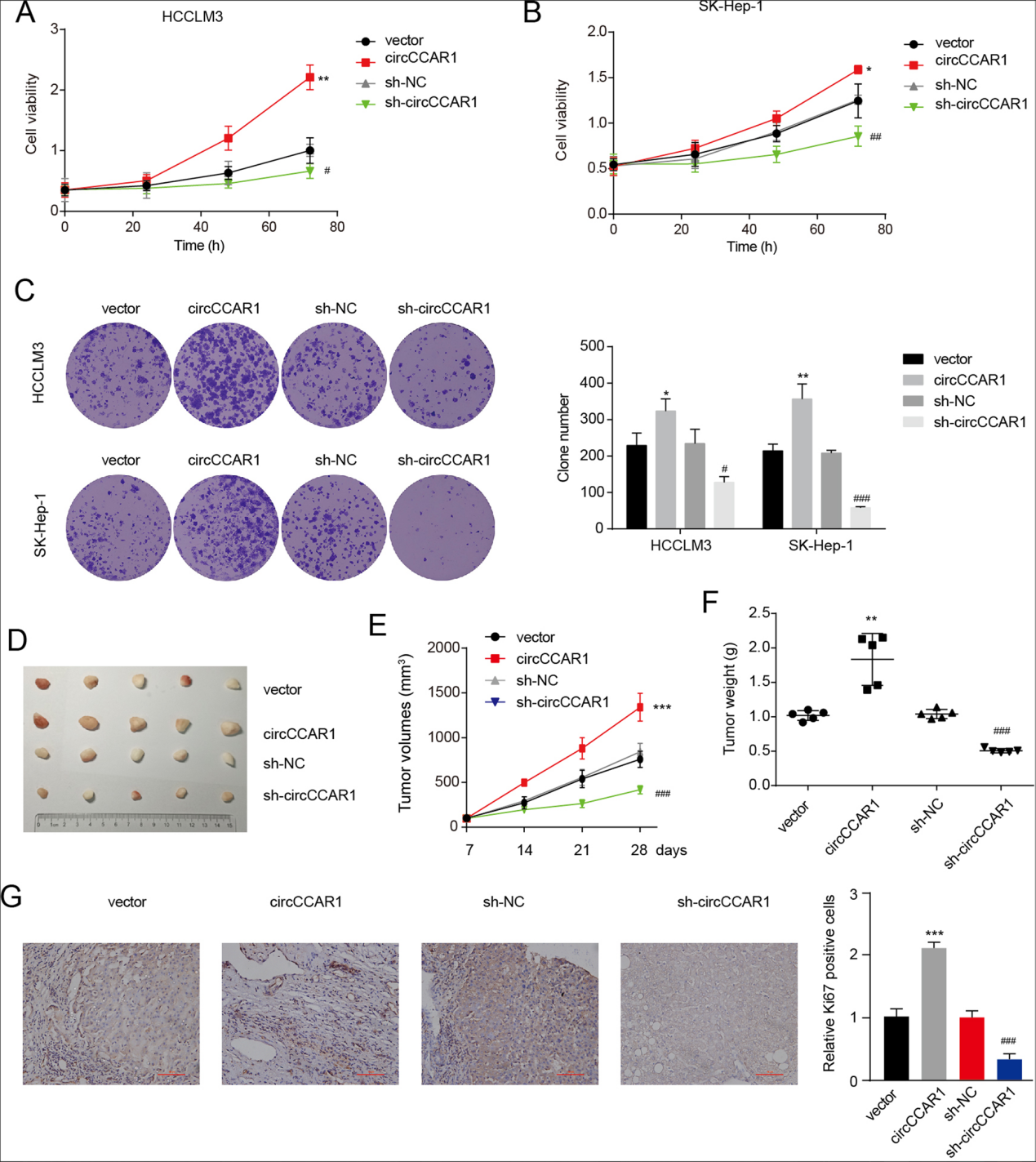
图2 . A-B.在circCCAR1过表达和敲低后的HCC细胞中进行CCK-8检测。C. 在circCCAR1过表达和敲低后的HCC细胞中进行集落形成实验。D. 异种移植肿瘤照片(n = 5)。E. 异种移植肿瘤生长曲线。F. 测定肿瘤重量。G. 异种移植瘤的GKi6。
结论3
circCCAR1在体外和体内促进HCC转移。
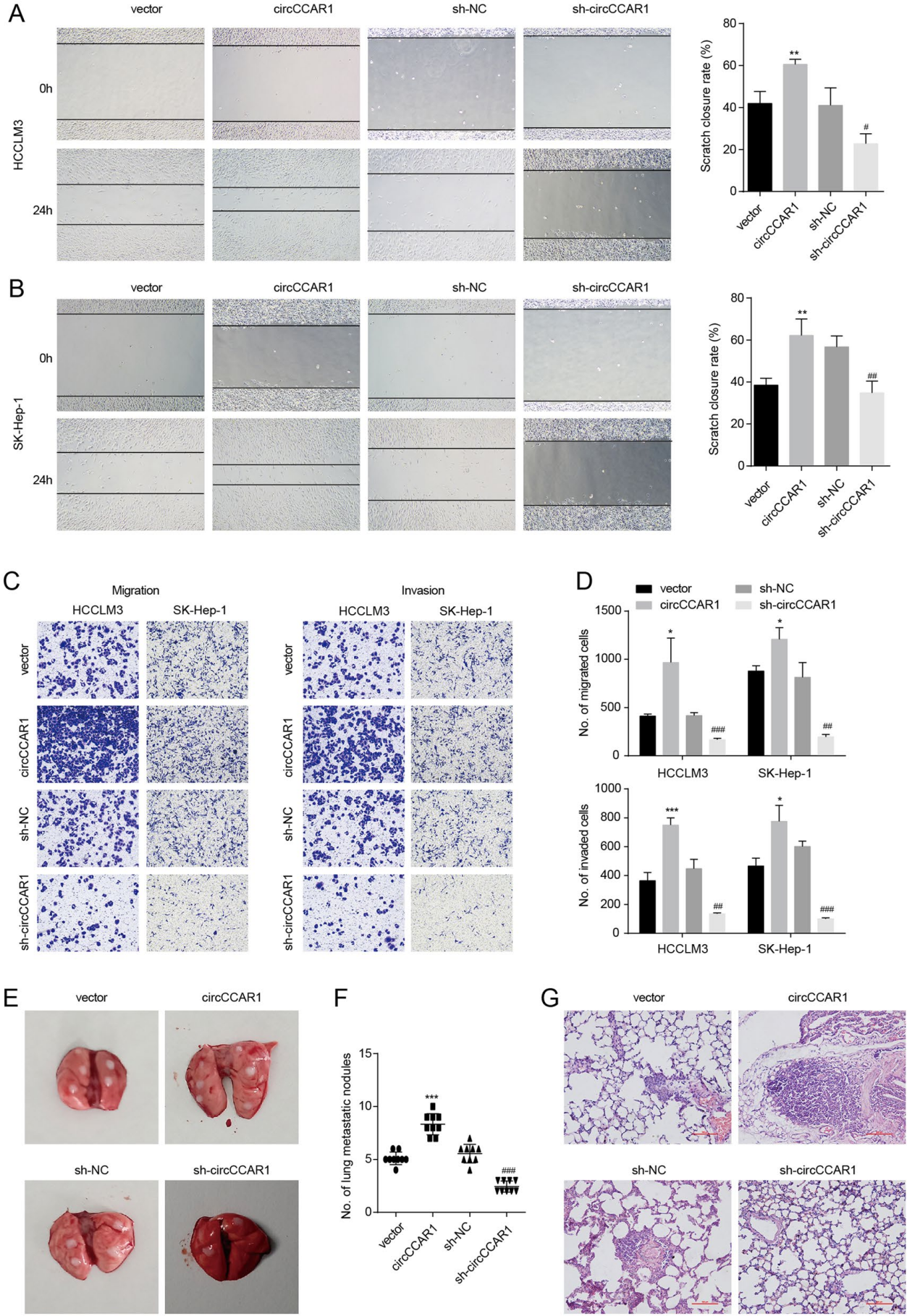
图3. A-B. 通过伤口愈合试验评估HCC细胞的迁移能力。C-D. 采用Transwell法评估HCC细胞的迁移和侵袭能力。E. 小鼠肺转移的肉眼观察(n = 9)。F. 计算转移结节的数量。G. 肺切片进行H&E染色。
结论4
EP300敲低抑制circCCAR1的表达。
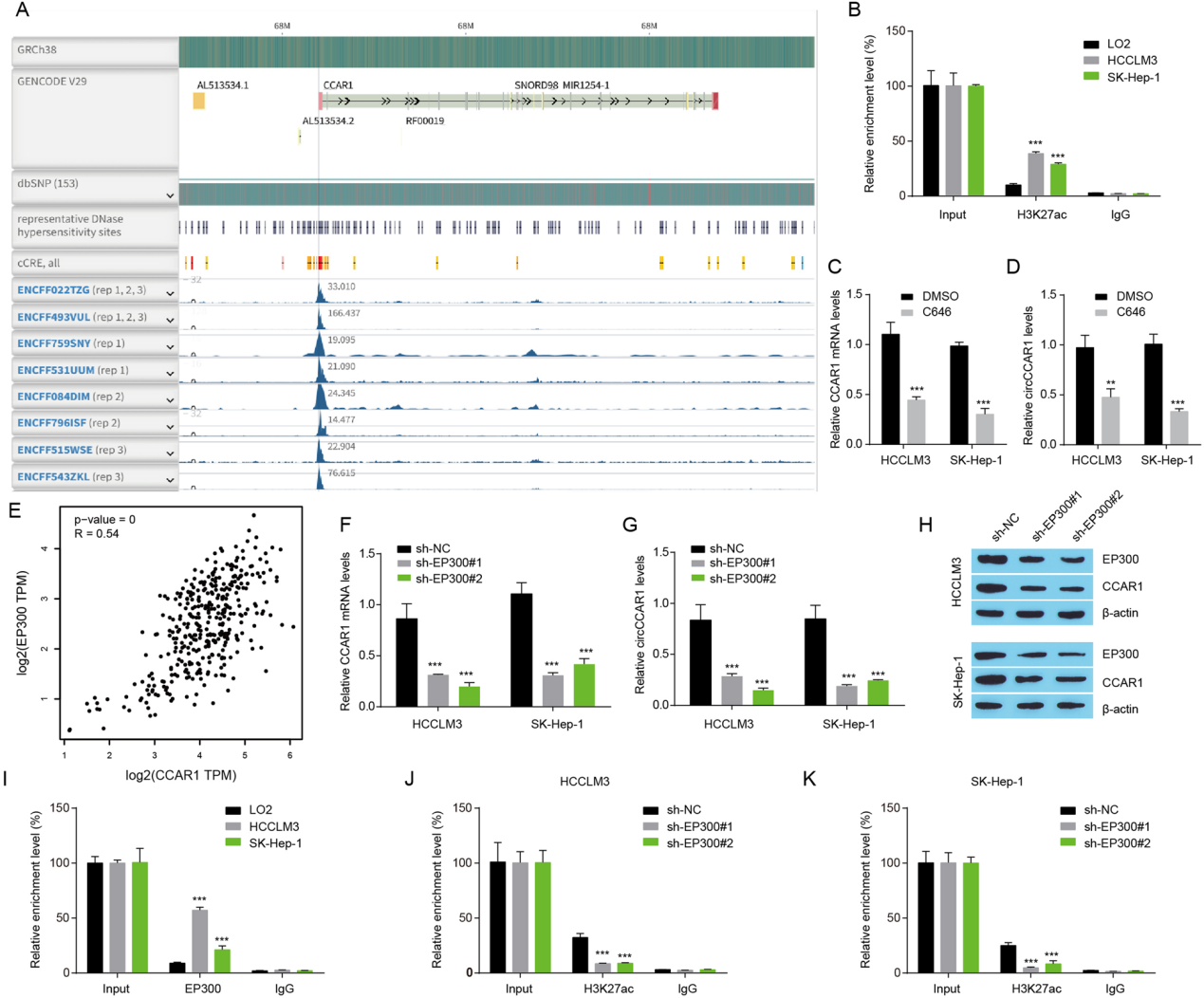
图4. A.分析HepG2细胞CCAR1启动子区。B. 在LO2、HCCLM3和SK-Hep-1细胞中,采用ChIP-qPCR检测CCAR1启动子中的H3K27ac信号。C-D. 采用qRT-PCR检测C646 (10 μM)或DMSO处理肝癌细胞48 h后CCAR1 mRNA和circCCAR1的表达。E. TCGA肝细胞癌中EP300与CCAR1的相关性分析。F‑G. CCAR1 mRNA和EP300缺失后检测circCCAR1的表达。H. EP300缺失后HCC细胞中EP300和CCAR1蛋白水平。I. 在LO2、HCCLM3和SK-Hep-1细胞中,采用ChIP-qPCR检测CCAR1启动子中的EP300信号。J-K. 在EP300沉默的HCC细胞中,使用ChIP-qPCR检测CCAR1启动子中的H3K27ac信号。
结论5
EIF4A3促进circCCAR1的环化和细胞质输出。
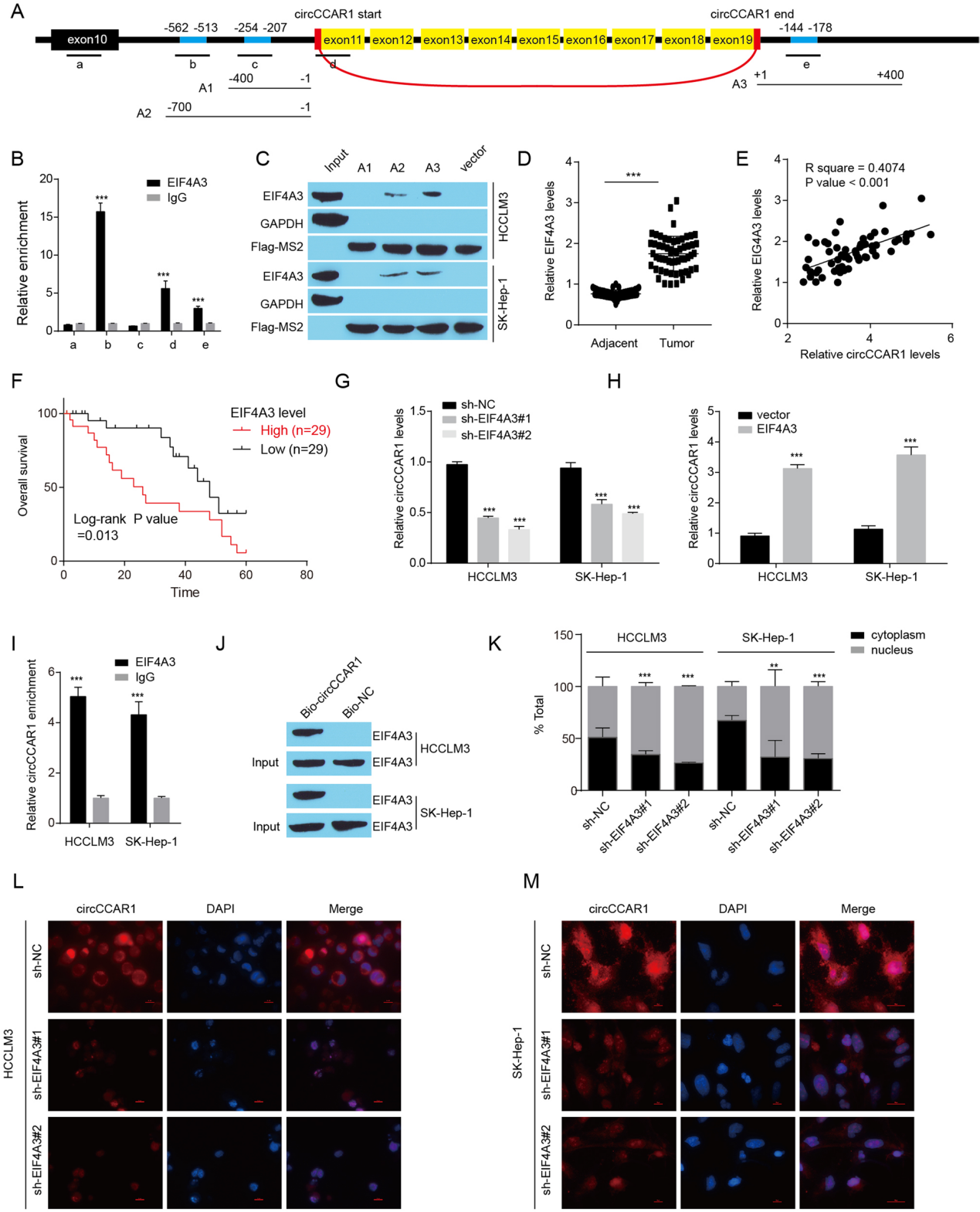
图5. A. 选择circCCAR1 pre-mRNA中的5个位点(a -e)设计qPCR引物,构建含有EIF4A3结合位点的3个质粒(A1-A3),用于EIF4A3蛋白pulldown实验。B. 通过RIP实验证实EIF4A3与circCCAR1 pre-mRNA之间存在相互作用。C. 检测EIF4A3蛋白MS2-RNA下拉复合体。D. 检测肝癌组织及其癌旁组织中EIF4A3水平。E. 计算肝癌样本中EIF4A3与circCCAR1水平的关系。F. 肝癌患者中EIF4A3表达与总生存率的关系。GH. EIF4A3缺失或过表达的HCCLM3和SK-Hep-1细胞中circCCAR1的表达量。I. 通过RIP实验证实EIF4A3与circCCAR1结合。J. 生物素化反义寡聚物pulldown测定EIF4A3蛋白水平。KM. 采用核细胞质分离法和FISH-IF法评估EIF4A3耗损后肝癌细胞中circCCAR1的细胞质输出。
结论6
WTAP介导的m6A修饰通过IGF2BP3增强了circCCAR1的稳定性。
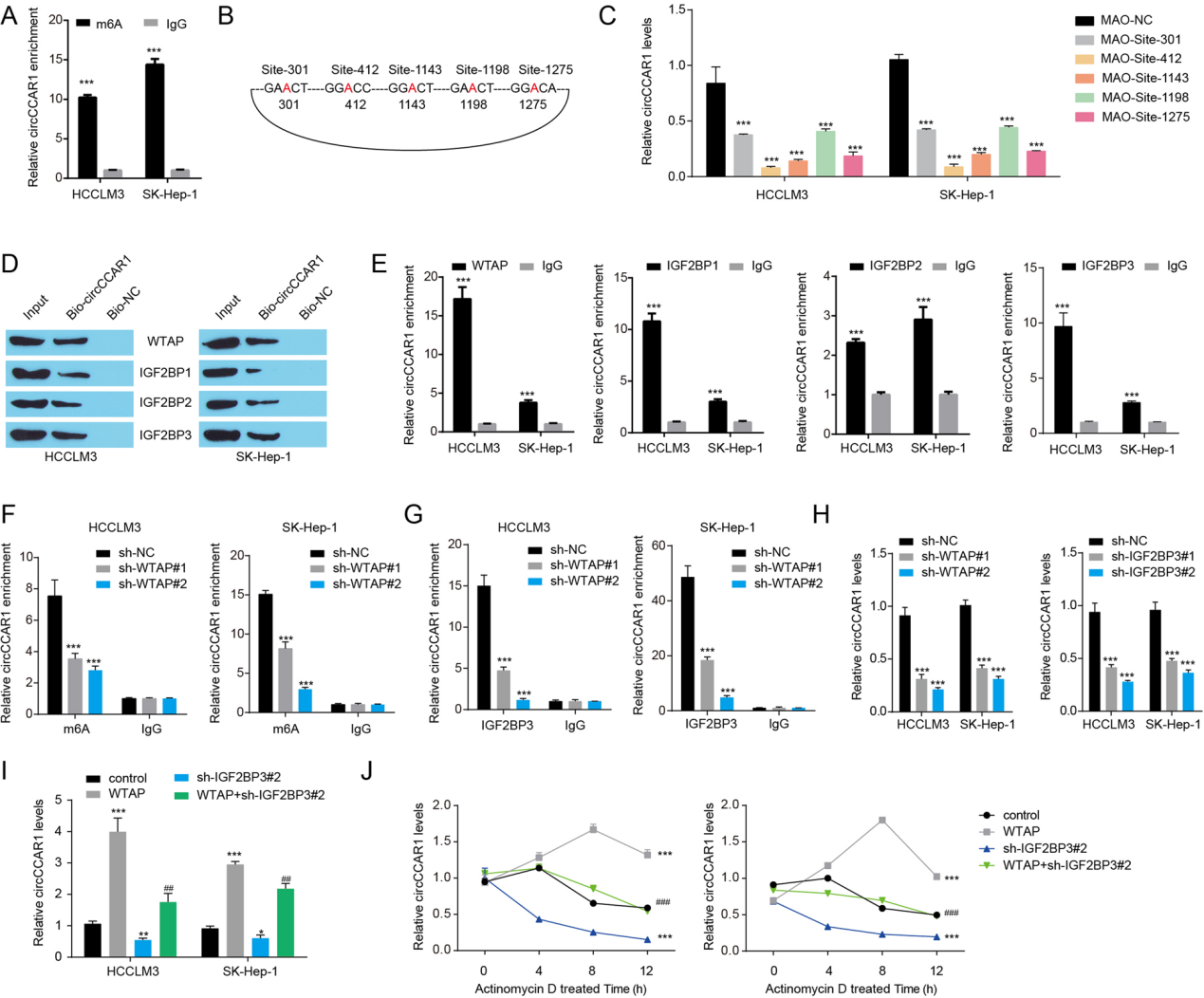
图6. A. m6A在circCCAR1的富集。B. m6A在circCCAR1中具有高综合得分的位点示意图。C. CircCCAR1在肝癌细胞中的表达,在MAO-circCCAR1 or MAO-NC 处理之后。D. 使用靶向circCCAR1环化位点的生物素标记的反义寡聚物进行WTAP和igfbp pulldown测定。E. RIP实验证实igf2bp与circCCAR1结合。F. 肝癌细胞中WTAP沉默后m6A在circCCAR1发生富集。G. 肝癌细胞WTAP敲除后circCCAR1中IGF2BP3的富集。H. WTAP敲低或IGF2BP3敲低后HCC细胞中的CircCCAR1水平。I. 肝癌细胞WTAP过表达或IGF2BP3敲低后circCCAR1表达水平。J. 放线菌素D处理下WTAP过表达或IGF2BP3敲低后肝癌细胞中circCCAR1的稳定性。
结论7
外泌体circCCAR1在HCC患者中升高。
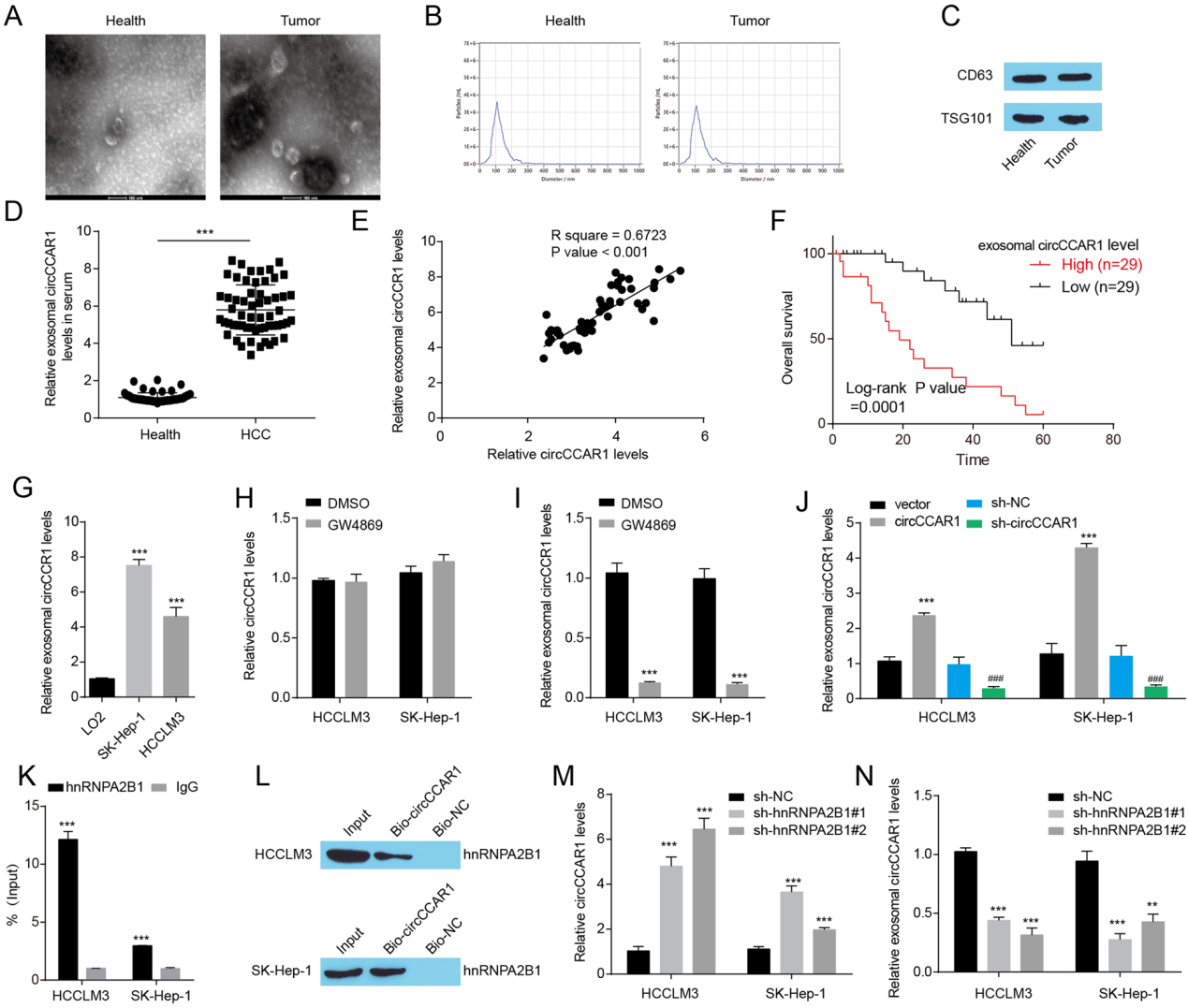
图7. A. 透射电镜检测外泌体。B. 外泌体大小分布和数量的NanoSight颗粒跟踪分析。C. 纯化外泌体中外泌体标志物(CD63和TSG101)的水平。D. 肝癌患者和健康供者血清外泌体中circCCAR1的水平。E. 采用Spearman相关分析评估血清外泌体circCCAR1与肿瘤样本circCCAR1的相关性。F. 肝癌患者外泌体circCCAR1水平与总生存率的关系。G. LO2细胞和HCC细胞培养液中外泌体中circCCAR1的水平。HI. 用GW4869处理HCC细胞的上清液分离的HCC细胞或外泌体中circCCAR1的水平。J. HCC细胞中circCCAR1过表达或缺失后外泌体中circCCAR1的水平。K. RIP实验证实circCCAR1与hnRNPA2B1结合。L. 通过RNA拉下实验证实circCCAR1与hnRNPA2B1结合。M. 敲低hnRNPA2B1后HCC细胞中circCCAR1的相对表达。N. 敲低hnRNPA2B1后外泌体中circCCAR1的相对表达。
结论8
外泌体circCCAR1通过稳定PD-1保护HCC细胞免受CD8+ T细胞的攻击。
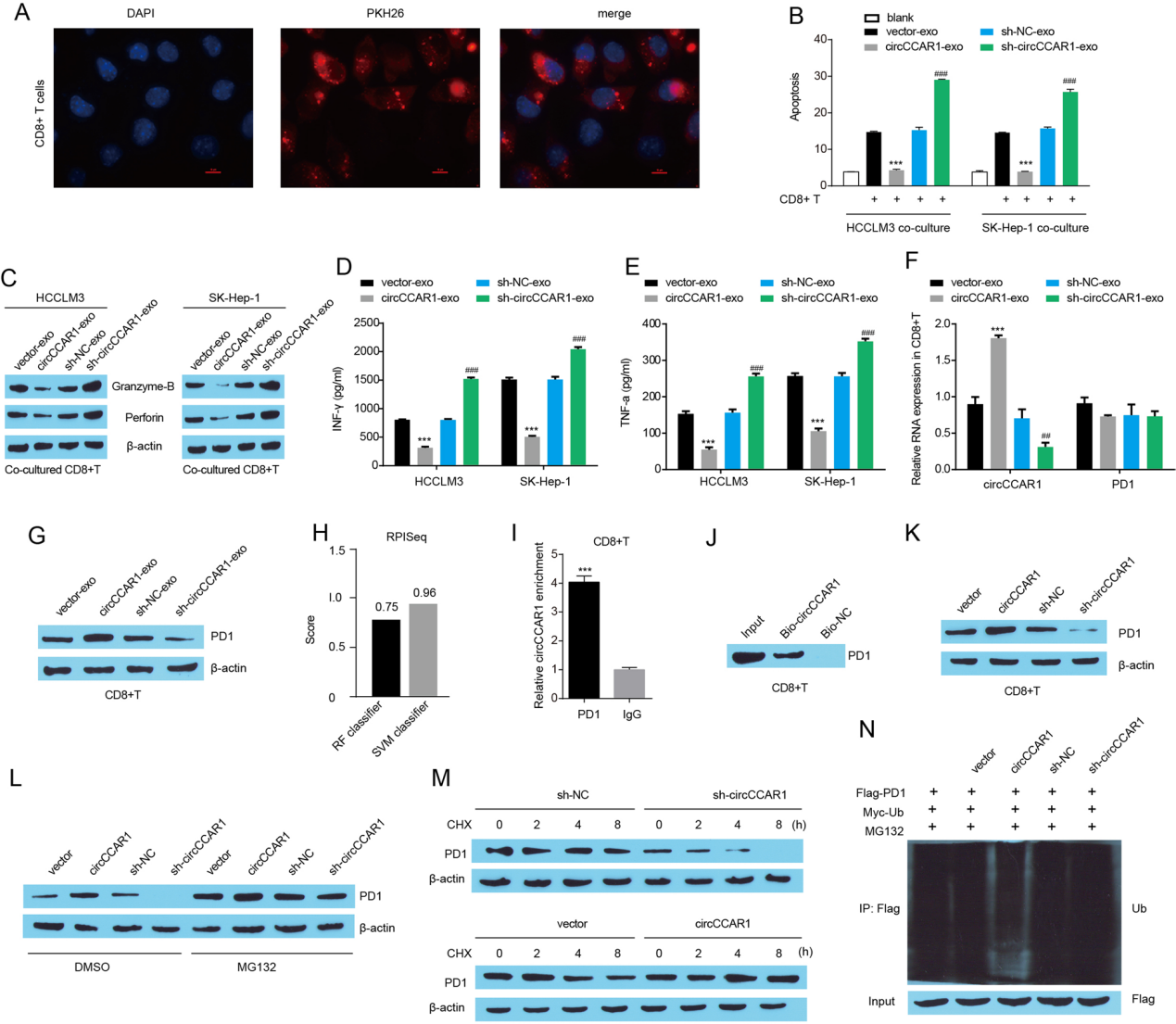
图8. A.CD8+T细胞内化PKH67标记的HCCLM3外泌体(红色)。B. CD8 + T细胞预处理的外泌体与HCC细胞共培养,然后通过FACS分析确定CD8 + T细胞介导的HCC细胞消除。CCD8 + T细胞穿孔素和颗粒酶B的水平。D-E. ELISA法检测CD8 + T细胞分泌的IFN-γ和TNF-α。F. circCCAR1和PD1 mRNA在CD8 + T细胞中的表达。G. CD8 + T细胞中PD1蛋白的表达。H. 利用RPISeq程序测定circCCAR1与PD1的相互作用强度。I. RIP实验证实circCCAR1与PD1结合。J. 使用生物素化的反义寡聚物靶向CD8 + T细胞中circCCAR1的结合位点,pulldown实验测定PD1水平。K. circCCAR1缺失或过表达后CD8 + T细胞中PD1的表达。L. 在circCCAR1缺失或过表达后,MG132处理CD8 + T细胞中的PD1蛋白水平。M. CD8 + T细胞经CHX处理后circCCAR1缺失或过表达后PD1蛋白的稳定性分析。N. CD8 + T细胞中PD1的泛素化测定。
结论9
circCCAR1促进HCC对抗PD1治疗的耐药性。
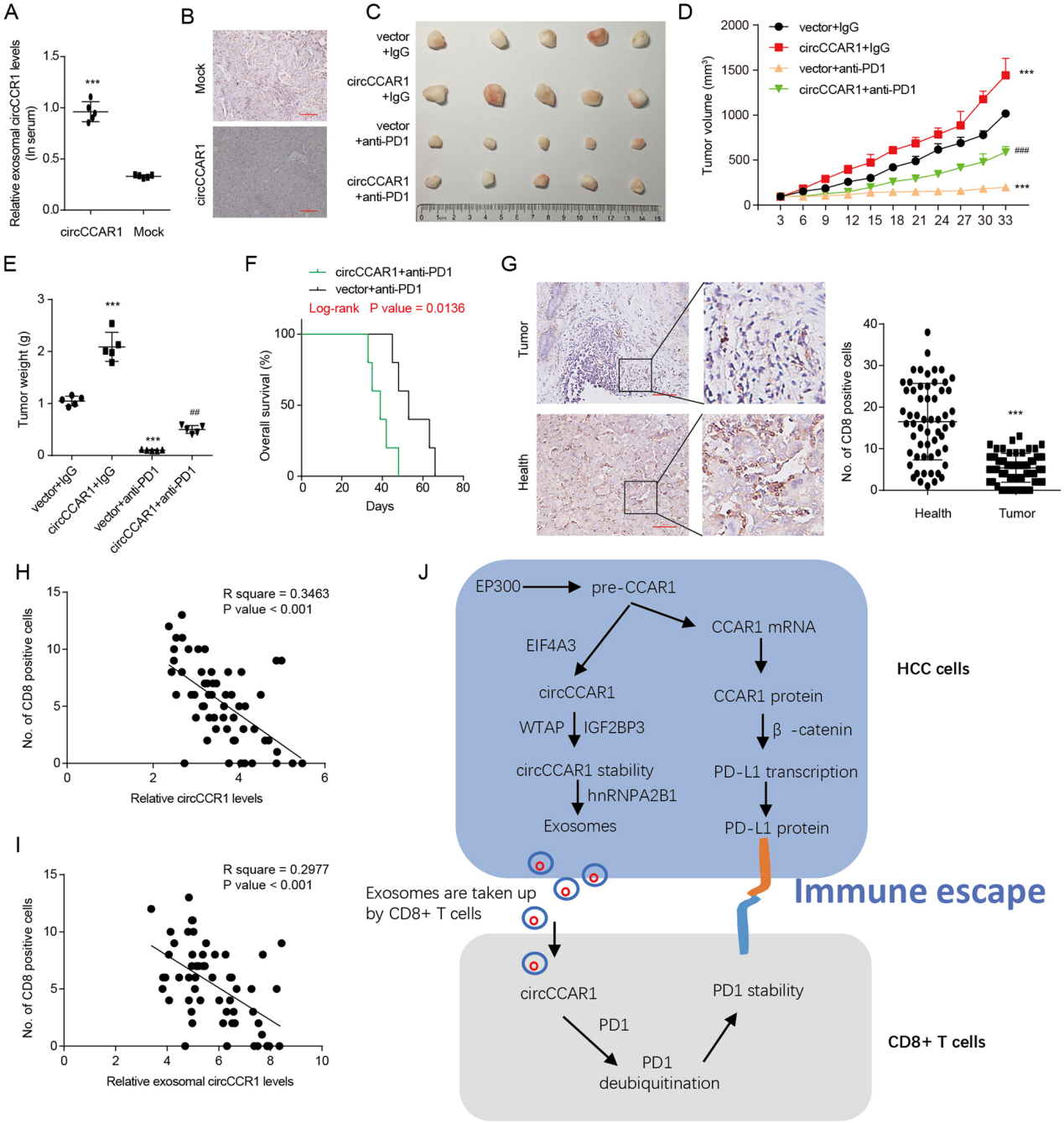
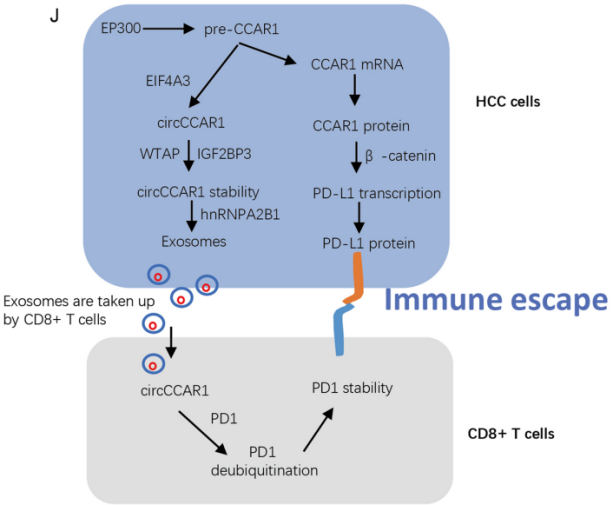
图9. A. HuNSG小鼠血清外泌体circCCAR1水平。B. 染色检测HCCLM3-circCCAR1或hcclm3 -模拟细胞来源的异种移植物中CD8阳性细胞。CE. 肿瘤体积和重量。F. 异种移植小鼠的存活曲线。G. 用CD8染色法分析HCC患者组织中CD8的含量。H. 计算circCCAR1与HCC组织中CD8阳性细胞的关系。I. 外泌体circCCAR1与基因表达的关系。J. 肝癌中circCCAR1介导的免疫抑制作用模型。
总结
综上所述,circCCAR1和CCAR1可能是肝癌患者的预后因素。circCCAR1/miR-127-5p/WTAP正反馈通路促进肝癌的生长和转移。肝癌细胞分泌的外泌体circCCAR1可以被CD8 + T细胞摄取完成激活,通过增强PD1的稳定性来促进细胞功能障碍。此外,肝癌细胞中CCAR1蛋白表达的增加通过结合β-catenin促进PD-L1的转录,这可能增强抗PD1治疗的耐药性。因此,靶向外泌体circCCAR1或CCAR1可能作为一种新的肝癌免疫治疗策略。
*本文系转载,如涉及版权等问题,请联系我们以便处理
 电话:400-9933-062
电话:400-9933-062 电子邮箱:business@wykt.com
电子邮箱:business@wykt.com

